 Print Article
Print Article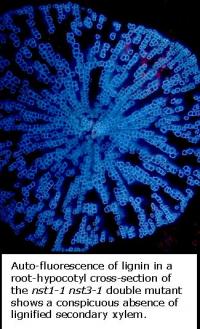
Auto-fluorescence of lignin in a root-hypocotyl cross-section of the nst1-1 nst3-1 double mutant shows a conspicuous absence of lignified secondary xylem. Wood is formed from secondary xylem tissue consisting of cells with a heavily thickened secondary cell wall that is enriched in lignin and cellulose. In contrast, primary cell walls are composed mainly of polysaccharides (including cellulose), but contain very little lignin. Wood formation occurs through a complex series of steps involving cell division and expansion and the biosynthesis of lignin and cellulose. In addition, heartwood (darker wood at the center of the trunk) forms in many tree species through a highly regulated process of programmed cell death.
Although herbaceous plants by definition do not form wood as in trees and shrubs, they nonetheless form secondary xylem tissue that is in many respects similar to that of their woody relatives. In two separate studies published in The Plant Cell, researchers identify key transcription factors that control the expression of genes related to secondary wall formation in the model herbaceous plant Arabidopsis. The presence of similar genes and pathways in tree species suggests that they may play a role in the regulation of wood formation in trees.
In the first paper, Masaru Ohme-Takagi and colleagues at the National Institute of Advanced Industrial Science and Technology in Japan, the Japan Science and Technology Agency, Nagoya University, and the RIKEN Institute (Mitsuda et al., 2007) show that two plant-specific transcription factors, designated NAC SECONDARY WALL THICKENINGS PROMOTING FACTOR1 (NST1) and NST3, regulate the formation of secondary walls in woody tissues (e.g. stem and hypocotyl) of Arabidopsis. Experiments with both loss- and gain-of-function mutants of these genes suggest that NST1 and NST3 function redundantly to promote secondary wall thickening in xylem tissue. Mutations disrupting the function of NST1 or NST3 alone did not produce any obvious abnormalities in mutant plants, but double knock-out lines of NST1 and NST3 showed a significant loss of thickening and lignification of secondary walls in stem and hypocotyl tissue. The double mutant plants were not able to remain upright and stems were easily bent and broken. This defective phenotype could be almost completely reversed by introducing into the double mutant a genomic fragment containing either the fully functional NST1 or NST3 gene. This shows that these genes have an overlapping or redundant function in promoting secondary wall thickening in woody tissue (e.g. stems). Further examination of gene expression in the mutant lines, and in plant lines engineered to overexpress NST1 and NST3, showed that these transcription factors regulate the expression of genes involved in biosynthesis of secondary walls. This work confirms and extends observations previously reported by Mitsuda et al. (2005. The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell 17: 2993-3006) and by Zhong et al. (2006. SND1, a NAC Domain Transcription Factor, Is a Key Regulator of Secondary Wall Synthesis in Fibers of Arabidopsis. Plant Cell 18: 3158-3170). (In the study by Zhong et al. (2006), SND1 is the same protein referred to as NST3 in the current paper by Mitsuda et al.). Closely related genes (putative orthologs) to NST1 and NST3 are present in the poplar genome, suggesting that they might also function as key regulators of secondary wall formation (and therefore wood) in trees.
Secondary wall thickenings in plants are found not only in xylem tissue but also in seedpods and anthers (the pollen-bearing organs in the flower). It is the organization of the thickened walls in these tissues that leads to the splitting (dehiscence) of mature seedpods and anthers that releases seed and pollen, respectively, to the atmosphere. In the second paper, Zoe A. Wilson and coworkers at the University of Nottingham, U.K. (Yang et al., 2007) show that the transcription factor MYB26 regulates secondary wall thickening in anthers of Arabidopsis. The authors make use of a male-sterile mutant of Arabidopsis, in which sterility is due to a failure of anther dehiscence (and consequent failure to release pollen) resulting from an absence of secondary wall thickening in anther tissue. As the anther dehydrates, the lack of secondary wall thickening does not allow the development of shearing forces necessary for the anther to split open, or dehisce. This defect previously was shown to be due to loss of function of the transcription factor MYB26 (Steiner-Lange et al. 2003. Disruption of Arabidopsis thaliana MYB26 results in male sterility due to nondehiscent anthers. Plant J. 34: 519–528). Yang et al. show that MYB26 regulates secondary wall thickening by controlling the expression of NST1 and the related gene NST2, which in turn regulate the expression of other genes linked to secondary thickening.
Results from both studies suggest that NST1 and NST2 function together in the regulation of secondary wall thickening in the anther, whereas NST1 and NST3 coordinately regulate secondary wall thickening in xylem tissue. MYB26 was found to influence the expression of NST1 and NST2 in relation to function in the anthers, and it is suggested that MYB26, or a related MYB protein, could also influence secondary wall formation in xylem (and therefore wood formation in trees) because of its effect on NST1. Further study of these genes in woody species could be important for understanding wood formation, with possible utility in genetic engineering of wood, paper, and biofuel crops. Moreover, these studies demonstrate that the herbaceous plant Arabidopsis can also serve as a model for woody species. Source : American Society of Plant Biologists
 Mail to a Friend
Mail to a Friend
